CTLA-4抑制剂可以全面协同PD-1抑制剂,增强免疫系统抗癌效果
癌症这个“大魔头”似乎知道,James Allison是它的克星。
它先是在Allison十一岁的时候,派淋巴瘤夺走了Allison的母亲[1]。随后,Allison的一位兄弟和两位叔叔也因癌症死亡[2]。亲人的离去,没有击垮Allison,反而促使他开始思考一个困扰了科学家很久的问题:免疫系统究竟为什么不能像消灭感冒那样消灭癌症?
历经30余年的探索,他带领团队破解了T细胞识别并杀死癌细胞的秘密,最终促成癌症免疫治疗的诞生。就在此时,癌症对Allison起了杀心,在短短12年内,先后派出前列腺癌、黑色素瘤和膀胱癌暗杀Allison[3],结果却惨遭反杀。

▲ 抗CTLA-4抗体和抗PD-1抗体治疗癌症获得2018年诺奖
2018年,Allison和Tasuku Honjo因开创癌症免疫治疗获得诺奖,是免疫学混沌100年的终结,也是免疫治疗攻克癌症时代的开始。
百年谜题终获破解,免疫治疗时代来临
在免疫治疗诞生之前,手术的使用已经有3000年的历史,放疗有120余年的历史,化疗也有70余年的历史。
这三种传统治疗方法被誉为癌症治疗的“三驾马车”。但是这三种方法仅对不到一半的癌症患者有效,还有另一大半患者不能上车。
逐渐地,科学家们认识到,散布于人体各处的、数以亿计的免疫细胞,形成了一个高效的防御系统。它们搜寻并消灭侵入人体的病原体,以及人体内衰老或其他异常的有害细胞。
直到1973年,也就是Allison博士毕业的那一年,科学家才终于认全了巨噬细胞、B细胞、T细胞、自然杀伤细胞和树突状细胞。现代免疫学的新纪元自此拉开帷幕。今年9月10日,美国科学家Max Cooper和澳大利亚科学家Jacques Miller,也因为发现B淋巴细胞和T淋巴细胞,获得有诺贝尔奖“风向标”之称的拉斯克奖。

▲ 拉斯克奖官网
不过,在当时科学家们仍然不知道免疫细胞是如何识别并杀死癌细胞的,更不知道为什么免疫细胞会对癌细胞视而不见。
后来,科学家在T细胞表面发现了T细胞独有的抗原识别受体(TCR)[4-7]。T细胞识别癌细胞的的方式总算被找到了,但是决定T细胞是否杀死癌细胞的那个关键分子还没找到。虽然后来发现了CD28分子[8],但是Allison等科学家发现,CD28分子应该也不是起关键作用的那个分子。
Allison的博士后Matthew Krummel很快就找到了头绪。决定T细胞是否抗癌的关键分子可能是法国科学家团队于1987年克隆的CTLA-4[9]。1991年,百时美施贵宝的Peter Linsley证实了这一发现[10]。而且,一年之后Linsley率先发表了抗CTLA-4抗体的研究成果,他认为CD28和CTLA-4协同作用,促进T细胞的增殖[11]。
尽管如此,Allison并不相信Linsley完全解决了T细胞的活化问题。在Allison看来,要想让车跑起来,有两种方法,一种是踩油门,一种是松刹车。大家都把CTLA-4当油门研究,它有没有可能是个刹车呢?
还真是。
经过两三年的苦战,到1995年Krummel终于证实了Allison的猜想:CTLA-4的作用与T细胞受体完全相反,它其实是个T细胞活性的抑制分子,也就是说CTLA-4是个“刹车”分子[12]。芝加哥大学的Jeffrey Bluestone团队也发现了这一点[13,14]。

▲ 抗CTLA-4抗体松手刹(诺贝尔奖官网)
这个发现让科学家感到意外,毕竟之前几乎没人想过T细胞上还会有刹车分子。决定命运的关键时刻又到了:这个CTLA-4分子究竟能为人类的健康做点儿什么呢?
Allison把这个新研究课题交给了研究过肿瘤的博士后Dana Leach。
Leach按照Allison的试验设计,把一批携带肿瘤的小鼠分成两组,一组注射抗CTLA-4抗体,另一组作为对照。最终的结果让Allison有点儿不敢相信:用了抗CTLA-4抗体的小鼠都活着,而且肿瘤都消失;而对照组小鼠,肿瘤一直疯长,最终所有的小鼠都死亡。
为了保险起见,他们又重新做了一遍实验。而且,这一次是Allison亲自动手。由于知道这个研究的价值和意义,Allison每天都会去看看鼠笼里的小鼠,并认真记录小鼠肿瘤的大小。
让Allison感到失望的是,在接连好多天的时间里,所有小鼠的肿瘤都在疯长,根本没有被抑制的迹象。几天之后,就在Allison打算放弃的时候,奇迹出现了:有两笼小鼠的肿瘤体积在缩小,另外两笼仍在长大。
原来阻断CTLA-4可以抗癌。
1996年,Leach和Allison的这项重要研究成果被顶级期刊《科学》刊登[15]。

▲ 1996年的论文
有了这项研究成果,Allison结合其他研究团队的发现,对CTLA-4的作用机制做了个复盘。
Allison推测,CTLA-4作为一个免疫抑制因子,它可以阻止免疫系统攻击正常的组织[16-18];而癌细胞恰恰利用了内置于T细胞之中的这种免疫抑制机制,抑制了T细胞的活性,为自己的生存和发展壮大提供了机会。
这时Allison有了个大胆的猜想,既然抗CTLA-4抗体可以治愈小鼠的癌症,或许在人身上也可以。
2000年,抗CTLA-4抗体终于在晚期黑色素瘤患者身上,向世人展示了免疫治疗的抗癌实力。2004年,第一个抗CTLA-4抗体的安慰剂控制,随机III期临床研究拉开帷幕,证实了抗CTLA-4抗体的强大治疗效果[19]。2011年,抗CTLA-4抗体成为全球第一个获批上市的免疫检查点抑制剂,标志着癌症的治疗进入了崭新的免疫治疗时代。
抗CTLA-4抗体的成功,让科研人员意识到,与CTLA-4类似的靶点肯定也有强大的抗癌潜力。
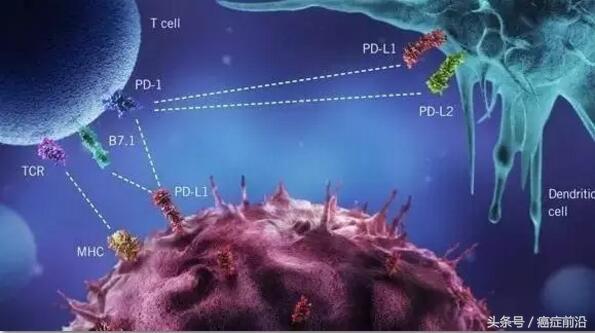
很快,日本京都大学Tasuku Honjo团队在1992年克隆到的PD-1分子吸引无数科学家的目光[20]。华人免疫学家陈列平和Tasuku Honjo的研究都表明,这个PD-1分子一旦与肿瘤细胞上的PD-L1结合,T细胞的抗肿瘤活性就没了[21-23]。这就意味着,PD-1和CTLA-4类似,也是T细胞的一种负调节因子。
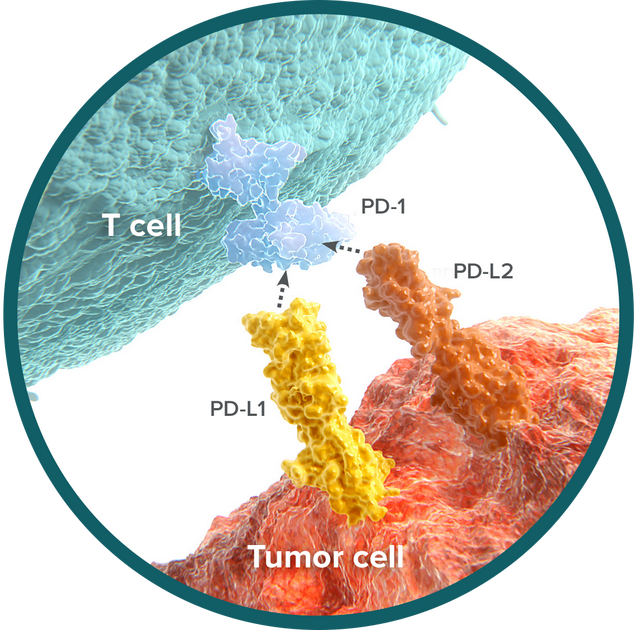
▲ PD-1通路
2014年,两款抗PD-1抗体先后获得FDA的批准上市,癌症免疫治疗的时代全面到来。
独立通路强强联合,免疫治疗再攀高峰
在这个崭新而充满希望的时代,我们一次次被免疫治疗的神奇效果折服。
我们已经看到,22%的晚期黑色素瘤患者,在接受抗CTLA-4抗体治疗之后,存活时间超过10年[19,24]。靶向PD-1/PD-L1通路的抗体也取得了良好的治疗效果[25-27]。尤其是在非小细胞肺癌中,在今年的世界肺癌大会上,CheckMate 017/057的5年随访数据发布,接受抗PD-1抗体治疗的晚期非小细胞肺癌患者的5年生存率,是接受化疗患者的5倍以上。
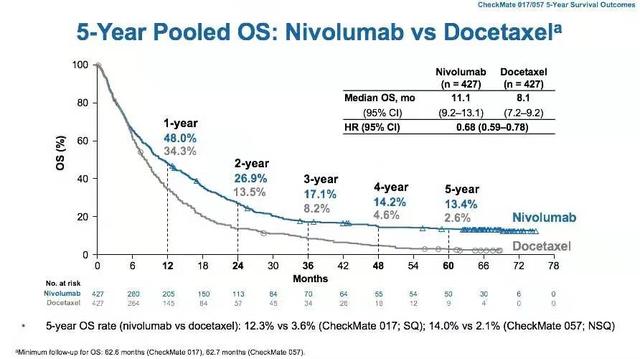
▲ CheckMate 017/057的5年随访数据
在兴奋之余,科学家和医生们都注意到,免疫治疗的神奇效果,也只能拯救30%左右的晚期癌症患者,仍有很大一部分患者不能从免疫治疗中获益。
有些科学家开始思考,如果把抗CTLA-4抗体和抗PD-1抗体联合使用,会不会解决一部分问题呢?
要解决这个问题,首先需要弄清楚的是抗CTLA-4抗体和抗PD-1抗体的作用部位和机制。
我们先来看看CTLA-4。通过前面的介绍,我们已经知道CTLA-4它是个“刹车”分子。它发挥“刹车”作用的过程也并不复杂。
我们身体在遭遇病原物或者异常细胞侵袭时,抗原呈递细胞会拿着它们的分子特征,进入淋巴结,激活初始T细胞,让T细胞开始扩充军队,抵抗外敌;在活化到一定程度之后,为了避免活化T细胞过多,而伤及健康的组织和细胞,初始T细胞就开始表达低水平的CTLA-4,刹车机制开始启动,“扩军”停止[31]。
此外,除了被激活的T细胞之外,有免疫抑制功能的调节T细胞也一直在大量表达CTLA-4。

▲ 抗PD-1抗体和抗CTLA-4抗体的作用机制并不相同[28]
再来看看PD-1。作为另一个“刹车”,PD-1与CTLA-4非常不同。首先,淋巴结里面的初始T细胞是不表达PD-1的,但是活化后的、杀伤力比较大的T细胞PD-1表达水平非常高。为了避免被这种T细胞误伤,人体大多数健康组织细胞都表达PD-1的配体PD-L1,只要PD-1见到PD-L1就认作朋友。
另外,有很多免疫细胞也表达PD-1,例如有免疫抑制功能的调节T细胞。
总结一下就是,抗CTLA-4抗体起作用的部位主要是淋巴结,它能解除CTLA-4对T细胞活化的抑制,增加外周血中活化T细胞的水平[29];它还能结合到调节T细胞表面。而抗PD-1抗体主要作用于外周血或者肿瘤中的被活化的T细胞,阻断这些T细胞的PD-1与免疫细胞的PD-L1结合,解除癌细胞对T细胞抗癌活性的抑制;它也能活化调节性T细胞。
那么抗CTLA-4抗体和抗PD-1抗体的不同作用机制,如何相互配合,提高抗癌效果的呢?这个咱们从近两年的研究成果说起。
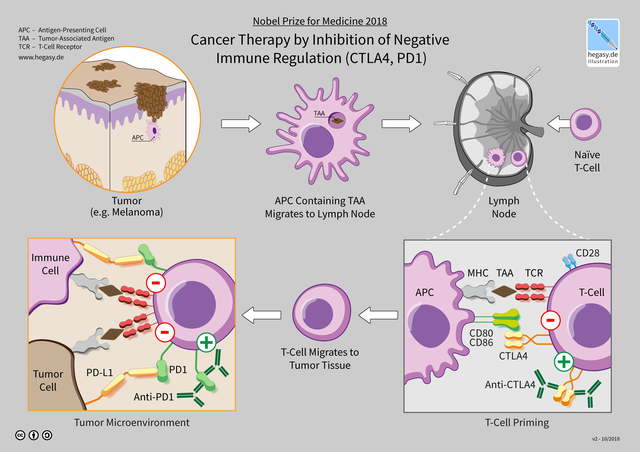
▲ 抗CTLA-4抗体和抗PD-1抗体作用机制(wiki)
大约在一年前,来自新加坡SIgN的研究团队发现了一个让人无奈的现实:肿瘤组织里面不仅有杀灭癌细胞杀伤性T细胞;还有大量与癌症无关的、原本应该是识别病毒的“旁观者”杀伤性T细胞,而这些T细胞根本就“不认识”癌细胞[30]。荷兰癌症研究所的科学家证实了SIgN的发现,而且进一步确认,肿瘤中具有识别癌细胞能力的杀伤性T细胞不足10%[31]。
更糟糕的是,即使是这10%的杀伤性T细胞,也不是抗PD-1抗体治疗之后的抗癌主力。就在两个月前,斯坦福大学医学院的科学家发现,那些在抗PD-1抗体治疗之前进入肿瘤,而且已经处于耗竭状态的肿瘤浸润T细胞,抗PD-1抗体拯救不了;而真正响应抗PD-1抗体治疗,最终起到抗癌作用的可能是治疗后从外周血进入肿瘤的T细胞[32]。
如果事实确实如此的话,那么抗CTLA-4抗体治疗提升外周血活化T细胞水平和记忆T细胞的能力,对抗PD-1抗体而言,是个非常大的助力。
二者联合使用的优势互补作用远不止这些。
疾病超进展也是影响抗PD-1抗体治疗效果的重要原因之一。前不久,韩国科学家发现,在接受抗PD-1抗体治疗的晚期非小细胞肺癌患者中,出现疾病超进展的患者占到18.9%[33];早在2017年的一项多癌种研究发现,甚至有9%的患者在接受抗PD-1抗体治疗之后,因疾病超进展死亡[34]。

▲ 图片来自Pixabay
今年4月份,来自日本国立癌症中心的科学家揭示了超进展背后的部分机制。他们发现,抗PD-1抗体影响的不只是杀敌的细胞毒性T细胞,也有抑制免疫的调节T细胞[35]。抗PD-1抗体能提升调节T细胞的增殖能力和免疫抑制能力,进而抑制了杀伤性T细胞的抗肿瘤活性,最终导致了肿瘤超进展。
前面我们已经介绍过,这个调节细胞非常特殊,它不仅能表达PD-1,还能表达CTLA-4。既然抗PD-1抗体激活了它,那抗CTLA-4抗体能不能消灭调节T细胞,抵消抗PD-1抗体对调节T细胞的激活作用呢?
还真可以,至少抗CTLA-4抗体ipilimumab可以。这主要是因为Ipilimumab的设计非常特殊,它是IgG1型抗体,具有很强的抗体依赖的细胞介导的细胞毒性作用(ADCC)作用[36]。这就意味着,这个抗CTLA-4抗体是可以杀死与它结合的调节T细胞的;由于初始T细胞的CTLA-4表达水平远远低于调节T细胞,所以能免遭杀身之祸[37,38]。也就是说,抗CTLA-4抗体有望降低抗PD-1抗体的负面作用。
▲ 免疫细胞(该图片由David Mark在Pixabay上发布)
实际上来,从临床数据来看,抗PD-1抗体联合有ADCC功能的抗CTLA-4抗体,确实能降低肿瘤超进展患者的比例[39]。
而且,有ADCC功能的抗CTLA-4抗体联合抗PD-1抗体,已经在黑色素瘤[40]、肾细胞癌[41]和非小细胞肺癌[42]中获得成功。
总的来说,以上这些出自顶级研究机构的研究都表明,抗CTLA-4抗体可以诱导新生抗肿瘤T细胞反应,抗PD-1抗体主要是恢复T细胞的抗肿瘤作用,这是这两种抗体作用机制之间的不同。
不过,二者的这种不同之中还有协同。具体来说,抗CTLA-4抗体可以让外周血中有更多的活化T细胞和记忆T细胞,抗PD-1抗体则可以解除肿瘤对这些T细胞抗癌活性的抑制;抗PD-1抗体有可能会激活调节性T细胞,而具有ADCC活性的抗CTLA-4抗体可以清除调节性T细胞;抗CTLA-4抗体还能导致肿瘤PD-L1代偿性过表达,抗PD-1抗体则能阻断PD-L1与PD-1的结合。正是这种协同作用,增强了免疫系统的抗癌效果。
最后,还有个不得不提的好消息是,就在今天,抗CTLA-4抗体联合抗PD-1抗体的CheckMate 227数据发表在《新英格兰医学期刊》上,再次证实两个单抗一线联合治疗非小细胞肺癌,可以让患者获得更长的总生存时间,而且这种获益与PD-L1表达水平无关[43]。
目前还有很多抗CTLA-4抗体联合抗PD-1抗体的临床研究正在进行之中。有了上述基础研究的支撑,以及已获成功的临床研究的证实。抗CTLA-4抗体联合抗PD-1抗体有望将免疫治疗推上另一个高峰。
* ipilimumab尚未在中国大陆上市
参考资料:
[1].https://www.washingtonpost.com/nation/2019/03/25/texas-scientist-was-called-foolish-arguing-immune-system-could-fight-cancer-then-he-won-nobel-prize/?noredirect=on
[2].https://www.washingtonpost.com/national/health-science/cancer-fighting-power-couple-tackles-mysteries-of-the-immune-system/2017/09/29/db52e514-810e-11e7-ab27-1a21a8e006ab_story.html
[3].https://www.tmc.edu/news/2018/11/a-new-texas-icon-james-allison-ph-d-wins-the-nobel/
[4].Allison J P, Mcintyre B W, Bloch D, et al. Tumor-specific antigen of murine T-lymphoma defined with monoclonal antibody.[J]. Journal of Immunology, 1982, 174(3): 2293-2300.
[5].Kappler J W, Kubo R T, Haskins K, et al. The mouse T cell receptor: Comparison of MHC-restricted receptors on two T cell hybridomas[J]. Cell, 1983, 34(3): 727-737.
[6].Chien Y, Becker D M, Lindsten T, et al. A third type of murine T-cell receptor gene[J]. Nature, 1984, 312(5989): 31-35.
[7].Yanagi Y, Chan A M, Chin B, et al. Analysis of cDNA clones specific for human T cells and the alpha and beta chains of the T-cell receptor heterodimer from a human T-cell line[J]. Proceedings of the National Academy of Sciences of the United States of America, 1985, 82(10): 3430-3434.
[8].Martin P J, Ledbetter J A, Morishita Y, et al. A 44 kilodalton cell surface homodimer regulates interleukin 2 production by activated human T lymphocytes.[J]. Journal of Immunology, 1986, 136(9): 3282-3287.
[9].Brunet J, Denizot F, Luciani M, et al. A new member of the immunoglobulin superfamily—CTLA-4[J]. Nature, 1987, 328(6127): 267-270.
[10].Linsley P S, Brady W, Urnes M, et al. CTLA-4 is a second receptor for the B cell activation antigen B7.[J]. Journal of Experimental Medicine, 1991, 174(3): 561-569.
[11].Linsley P S, Greene J, Tan P, et al. Coexpression and functional cooperation of CTLA-4 and CD28 on activated T lymphocytes.[J]. Journal of Experimental Medicine, 1992, 176(6): 1595-1604.
[12].Krummel M F, Allison J P. CD28 and CTLA-4 have opposing effects on the response of T cells to stimulation.[J]. Journal of Experimental Medicine, 1995, 182(2): 459-465.
[13].Walunas T L, Lenschow D J, Bakker C Y, et al. CTLA-4 can function as a negative regulator of T cell activation[J]. Immunity, 1994, 1(5): 405-413.
[14].Lenschow D J, Zeng Y, Thistlethwaite, et al. Long-term survival of xenogeneic pancreatic islet grafts induced by CTLA4lg[J]. Science, 1992, 257(5071): 789-792.
[15].Leach D R, Krummel M F, Allison J P, et al. Enhancement of Antitumor Immunity by CTLA-4 Blockade[J]. Science, 1996, 271(5256): 1734-1736.
[16].Waterhouse P, Penninger J M, Timms E, et al. Lymphoproliferative Disorders with Early Lethality in Mice Deficient in Ctla-4[J]. Science, 1995, 270(5238): 985-988.
[17].Tivol E A, Borriello F, Schweitzer A N, et al. Loss of CTLA-4 leads to massive lymphoproliferation and fatal multiorgan tissue destruction, revealing a critical negative regulatory role of CTLA-4[J]. Immunity, 1995, 3(5): 541-547.
[18].Chambers C A, Sullivan T J, Allison J P, et al. Lymphoproliferation in CTLA-4–Deficient Mice Is Mediated by Costimulation-Dependent Activation of CD4+ T Cells[J]. Immunity, 1997, 7(6): 885-895.
[19].Hodi F S, Mcdermott D F, Weber R W, et al. Improved Survival with Ipilimumab in Patients with Metastatic Melanoma[J]. The New England Journal of Medicine, 2010, 363(8): 711-723.
[20].Ishida Y, Agata Y, Shibahara K, et al. Induced expression of PD-1, a novel member of the immunoglobulin gene superfamily, upon programmed cell death.[J]. The EMBO Journal, 1992, 11(11): 3887-3895.
[21]. Nishimura H, Nose M, Hiai H, et al. Development of Lupus-like Autoimmune Diseases by Disruption of the PD-1 Gene Encoding an ITIM Motif-Carrying Immunoreceptor[J]. Immunity, 1999, 11(2): 141-151.
[22]. Dong H, Zhu G, Tamada K, et al. B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion.[J]. Nature Medicine, 1999, 5(12): 1365-1369.
[23]. Freeman G J, Long A J, Iwai Y, et al. Engagement of the Pd-1 Immunoinhibitory Receptor by a Novel B7 Family Member Leads to Negative Regulation of Lymphocyte Activation[J]. Journal of Experimental Medicine, 2000, 192(7): 1027-1034.
[24].Schadendorf D, Hodi F S, Robert C, et al. Pooled Analysis of Long-Term Survival Data From Phase II and Phase III Trials of Ipilimumab in Unresectable or Metastatic Melanoma[J]. Journal of Clinical Oncology, 2015, 33(17): 1889-1894.
[25].Brahmer J R, Tykodi S S, Chow L Q, et al. Safety and Activity of Anti–PD-L1 Antibody in Patients with Advanced Cancer[J]. The New England Journal of Medicine, 2012, 366(26): 2455-2465.
[26].Topalian S L, Hodi F S, Brahmer J R, et al. Safety, Activity, and Immune Correlates of Anti–PD-1 Antibody in Cancer[J]. The New England Journal of Medicine, 2012, 366(26): 2443-2454.
[27].Gettinger S N, Horn L, Jackman D M, et al. Five-Year Follow-Up of Nivolumab in Previously Treated Advanced Non–Small-Cell Lung Cancer: Results From the CA209-003 Study[J]. Journal of Clinical Oncology, 2018, 36(17): 1675-1684.
[28].Buchbinder E I, Desai A M. CTLA-4 and PD-1 Pathways: Similarities, Differences, and Implications of Their Inhibition[J]. American Journal of Clinical Oncology, 2016, 39(1): 98-106.
[29].Robert L, Tsoi J, Wang X, et al. CTLA4 Blockade Broadens the Peripheral T-Cell Receptor Repertoire[J]. Clinical Cancer Research, 2014, 20(9): 2424-2432.
[30].Simoni Y, Becht E, Fehlings M G, et al. Bystander CD8+ T cells are abundant and phenotypically distinct in human tumour infiltrates.[J]. Nature, 2018, 557(7706): 575-579.
[31].Scheper W, Kelderman S, Fanchi L, et al. Low and variable tumor reactivity of the intratumoral TCR repertoire in human cancers[J]. Nature Medicine, 2019, 25(1): 89-94.
[32].Li J, Byrne K T, Yan F, et al. Tumor cell-intrinsic factors underlie heterogeneity of immune cell infiltration and response to immunotherapy[J]. Immunity, 2018, 49(1): 178-193. e7.
[33].Kim C G, Kim K H, Pyo K H, et al. Hyperprogressive disease during PD-1/PD-L1 blockade in patients with non-small-cell lung cancer[J]. Annals of Oncology, 2019, 30(7): 1104-1113.
[34].Champiat S, Dercle L, Ammari S, et al. Hyperprogressive Disease Is a New Pattern of Progression in Cancer Patients Treated by Anti-PD-1/PD-L1[J]. Clinical Cancer Research, 2017, 23(8): 1920-1928.
[35].Kamada T, Togashi Y, Tay C, et al. PD-1+ regulatory T cells amplified by PD-1 blockade promote hyperprogression of cancer[J]. Proceedings of the National Academy of Sciences of the United States of America, 2019, 116(20).
[36].Salfeld J G. Isotype selection in antibody engineering[J]. Nature Biotechnology, 2007, 25(12): 1369-1372.
[37].Peggs K S, Quezada S A, Chambers C A, et al. Blockade of CTLA-4 on both effector and regulatory T cell compartments contributes to the antitumor activity of anti–CTLA-4 antibodies[J]. Journal of Experimental Medicine, 2009, 206(8): 1717-1725.
[38].Bulliard Y, Jolicoeur R, Windman M, et al. Activating Fc γ receptors contribute to the antitumor activities of immunoregulatory receptor-targeting antibodies[J]. Journal of Experimental Medicine, 2013, 210(9): 1685-1693.
[39].Act A, Larkin J, Sileni V C, et al. Combined nivolumab and ipilimumab or monotherapy in untreated Melanoma[J]. The New England Journal of Medicine, 2015, 373(1): 23-34.
[40].Wolchok J D, Chiarionsileni V, Gonzalez R, et al. Overall Survival with Combined Nivolumab and Ipilimumab in Advanced Melanoma[J]. The New England Journal of Medicine, 2017, 377(14): 1345-1356.
[41].Motzer R J, Tannir N M, Mcdermott D F, et al. Nivolumab plus Ipilimumab versus Sunitinib in advanced renal-cell carcinoma[J]. The New England Journal of Medicine, 2018, 378(14): 1277-1290.
[42].Hellmann M D, Ciuleanu T, Pluzanski A, et al. Nivolumab plus Ipilimumab in Lung Cancer with a High Tumor Mutational Burden[J]. The New England Journal of Medicine, 2018, 378(22): 2093-2104.
[43].https://www.nejm.org/doi/full/10.1056/NEJMoa1910231?query=featured_home